|
|
|
|
 Joe Wolfe, School of Physics, The University of New South Wales,
Sydney, and
Joe Wolfe, School of Physics, The University of New South Wales,
Sydney, andLow temperature damage in cells can be divided into damage produced by three effects: (i) low temperature per se; (ii) direct effects of freezing and (iii) indirect effects of freezing. Cryobiology (the study of life at low temperatures) and anhydrobiology (the study of life at low water contents) have some features in common. This is because, in environmental freezing, one of the major causes of damage is freezing induced dehydration. This essay gives an introduction to several types of cryobiological and anhydrobiological damage at the cellular level. It concentrates however on freezing- or desiccation-induced cellular dehydration, and on our biophysical research in this area and on related systems.
At the end we list references to the scientific papers summarised here. Much of physics underlying this essay comes from Wolfe, J. Bryant, G., (1999) Freezing, drying and/or vitrification of membrane-solute-water systems. Cryobiology, 39, 103-129.
Further, cryogenic temperatures (those near the boiling point of nitrogen) are not dangerous per se. At liquid nitrogen temperatures, almost all biochemistry and physiology is so slow that nothing (including damage!) happens at those temperatures. Cooling to cryogenic temperatures may kill, and warming up may kill---we'll discuss these later---but little damage occurs in the cryopreserved state.
Freezing, however, is often deadly. On the scale of organs, the formation of ice can cause mechanical damage by expansion, or rupture as pointed ice crystals grow through the tissue. Further, if the ice forms inside the cell, the cell almost always dies. There have been a few recent reports of cells surviving intracellular ice formation (IIF) in vitro, but IIF is usually taken by working cryobiologists as indicating cell death.
Ice is a poor solvent. When ice forms in an aqueous solution, most solutes are excluded from the ice, and remain in a concentrated unfrozen solution. So concentrations become very large and may be toxic. High concentrations of salts affect electrical or ionic interactions, including those that help stabilise the native state of enzymes. The unfolding and denaturation of enzymes is often irreversible. Further, ice and water interact differently with the surfaces of membranes and macromolecules, which is important because these interactions (via the surface tension of water or the hydrophobic effect) are also involved in maintaining the healthy state of the cell's internal anatomy or ultrastructure.
Before we go on to look at freezing damage, consider the question: How to avoid intracellular ice formation? It may be asked and answered at the physiological, ecological and medical levels. Keeping warm is the simplest answer, although this strategy is unavailable to many living things in most environments. Four other strategies are supercooling, freezing point depression, dehydration and vitrification,. These occur to varying extents in both the artificial process of cryopreservation and in environmental freezing. In all cases, the integrity of the cell's plasma membrane is critical. An intact membrane is necessary to prevent extracellular ice from nucleating intracellular ice. But an intact membrane is vital for other reasons too (e.g. maintaining the different compositions of intra- and extracellular solutions) and rupture of the membrane is also widely used as an indicator of cell death.
Figure: A very simplified sketch shows a cell
before and after the freezing of its
tissue or supporting medium. The elevated osmotic pressure causes a large
reduction in the aqueous volume of the cell.
Consequently, all of the non-aqueous components are brought into close
proximity. In this state, stacks of membranes begin to resemble lamellar
phases.
Many species of plants and animals, and especially their seeds and spores,
survive freezing temperatures by cellular dehydration. Dehydration increases
the osmotic pressure of the intracellular solution (the cytoplasm) which
depresses its freezing temperature and promotes vitrification---both inhibit
intracellular ice formation. Equilibration with ice at about -20��C or with an
atmosphere of about 80% relative humidity requires that a cellular interior
have a composition of about 10 osmolal. Osmolal means roughly
'osmotically effective moles of solute per kg of water'. The amount of
dehydration required to achieve this depends on the initial composition. If all
solutes were ideal, and if the initial composition were 1�osmolal (a typical
value for a plant cell, and a few times higher than that of most animal cells),
then only 10% of the initial intracellular water would remain. If the initial
composition were 2 osmolal, then 20% would remain. In practice, these
water contents are underestimates because the osmotic pressure of many
solutions increases more than linearly with concentration, and because of
colligative properties of the cellular ultrastructure. Dehydration itself can
cause damage, which we discuss later in some detail. The next figure shows
the relationship among several variables that depend upon the chemical
potential of water, which is a function of the freezing temperature or the
atmospheric humidity.
Figure: This figure allows conversion among
several variables related to aqueous solutions. The first scale shows, on a log
scale, the reduction in the chemical potential with respect to pure water at
atmospheric pressure. Dividing this by the molecular volume of liquid water
gives it the dimensions of pressure. To the extent that water is
incompressible, this quantity equals the osmotic pressure (P) minus the hydrostatic pressure (P). The negative
of this quantity is called the water potential (Slayter, 1967). The
second scale shows the equilibrium freezing temperature of a solution whose
water potential is given in the first scale. The next scale shows the
composition (in osmolal) of a solution which would produce an osmotic
pressure given by the first scale. The scale at the bottom shows the relative
humidity of an atmosphere equilibrated with water having the water
potential given in the first scale, or with ice at a temperature given by the
second.
Molecules in a liquid undergo random, Brownian motion. For freezing to
occur in a supercooled liquid, the diffusing molecules must spontaneously
form a small cluster (called a nucleus or embryo) of molecules that
temporarily has a structure similar to that of ice. In a supercooled liquid such
clusters form and dissipate rapidly. If however the cluster is larger than some
critical size, it becomes energetically favourable for other diffusing
molecules to join the structure, and it grows through the sample
(crystallising, or freezing). This process is called nucleation and crystal
growth. Nucleation can be either homogeneous (as described above, and in
the figure below) or heterogeneous, where an impurity (or the container
wall) forms a substrate upon which nuclei can grow.
Figure: A simplified sketch of the effects of
solutes on the nucleation of freezing. Water molecules and solutes are
represented by the symbols w and s respectively. The arrows represent
diffusion, and the length of the arrows indicates the speed of diffusion. The
large circles represent the critical nucleation radius. In (a), only water is
present. For a critical nucleus to form, the water molecules in the volume
represented by the circle must spontaneously arrange themselves into a
regular ice-like structure. If this regular lattice is larger than critical size,
then the crystal will grow. (b) shows the situation in the presence of solutes
(the solute:water molar ratio is 1:4). First, solutes increase the viscosity, so
diffusion is reduced (hence smaller arrows in (b) than (a)). Second, in order
to form a critical nucleus, a volume equal to or greater that the critical radius
must be completely free of solute molecules. This is not the case here. As the
concentration of solutes increases, this effect becomes stronger, further
reducing the chance of nucleation.
The probability of nucleation in a supercooled liquid depends on several
factors: it increases with the volume of the sample and the degree of
supercooling; it decreases with increasing solution concentration; and it also
increases in the presence of impurities that can act as heterogeneous nuclei.
A pure liquid in a small volume with no impurities can be supercooled a
long way below its equilibrium freezing point. Small volumes (microlitres)
of pure water, for example, can be cooled to about -40 �C.
At very low temperatures, the viscosity of a solution rises sharply, and
molecular diffusion is reduced. If cooling is fast, then the viscosity rises
rapidly, hindering nucleation. If cooling is sufficiently fast, the viscosity can
become so large that molecular diffusion is effectively halted, and the
probability of nuclei formation becomes negligible. The sample is then said
to be a glass or vitreous solid, and the process is called
vitrification. A glass is amorphous (unlike a crystal, it has
no long range order) but has the mechanical properties of a solid. A material
is said to be a glass if its viscosity reaches 1014�Pa.s (Franks,
1982). A glass is by definition in a state of very long-lived non-
equilibrium.
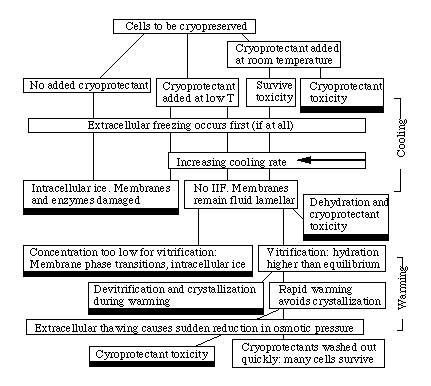
Figure: A simplified and incomplete flow chart
for cryopreservation showing some of the steps and some of the dangers.
The black rectangles represent cell death. Not shown on this chart is the
possibility of vitrification of small volumes in the absence of
cryoprotectants, using extremely rapid cooling.
The cytoplasm has a higher solute concentration, because of a combination
of dehydration and the addition of cryoprotectants that can permeate through
the membrane. This, together with the rapid cooling rate, may together allow
vitrification as discussed above. Higher cooling rates allow lower doses of
the toxic cryoprotectants, but cooling rates are often limited in practice by
heat conduction in the samples being cooled, particularly for macroscopic
organs.
Once the cell is vitrified, "suspended animation" is nearly achieved:
all processes of metabolism and injury are slowed to almost zero. Provided
that the sample avoids mechanical shocks (glasses are brittle!), the next
threat it faces is crystallisation during warming. The vitreous state is
unstable with respect to ice plus concentrated solution, but it is prevented
from achieving the stable state by its very high viscosity. As the temperature
rises, the viscosity falls, molecular motion becomes less slow, and water
molecules may diffuse and rotate into the configurations required to nucleate
ice, or to add to existing nuclei. The chance of large scale ice nucleation
and/or growth occurring depends on the time the sample is exposed to lower
viscosities while it is below the equilibrium freezing temperature. Thus
successful warming should cross this temperature range quickly (Rall et al.,
1984). As is the case with cooling, warming rates are usually limited by
conduction. Microwave heating has been proposed, but the rapidly changing
absorption spectrum makes this more difficult (Baudot, 1997). There is the
further problem that rapid but inhomogeneous heating can produce
dangerous mechanical stresses in macroscopic tissues.
Figure: A simplified flow chart for freeze-thaw
damage in some species that are capable of acclimation including solute
accumulation. The black rectangles represent cell death.
The required equilibrium dehydration can be substantial. In an earlier figure we saw that freezing to -10��C requires
osmotic pressures of more than 10�MPa and solutions of several osmolal.
For typical cells with osmolalities less than one, this requires a several-fold
reduction in water content. We are mainly concerned with the physical
stresses produced by this freezing-induced dehydration, but we shall briefly
describe two other damage mechanisms first.
A range of symptoms of damage have been reported in the severely
dehydrated state for both model membrane systems and the membranes of
living cells. (i)�In the lipids that are the major molecular component of cell
membranes, the gel-liquid crystal phase transition occurs at higher
temperatures (e.g. Crowe et al., 1988, Tsvetkov et al., 1989, Koster et al.,
1994 and references in these papers). This is important because coexistence
of the phases has been associated with reduced semipermeability.
(ii)�Membranes may undergo topological changes. Electron micrographs
show membranes associated with arrays of long cylinders which resemble
the inverted hexagonal phase formed by some lipid-water dispersions at very
low hydration. In this phase, the water is found in long narrow cylinders on a
hexagonal array, each cylinder surrounded by the hydrophilic moiety of the
lipids. This geometry renders them unsuitable for forming a semipermeable
barrier. Other topological changes have also been reported (Fujikawa 1995,
Gordon-Kamm and Steponkus 1984, Uemura et al., 1995, Webb et al.,
1993). (iii)�Lateral phase separations may occur in the fluid state. At low
hydrations, large areas of protein free membrane are observed in electron
micrographs. Further, phase separation may occur to produce one phase rich
in highly hydrating lipid species, and another rich in weakly hydrating
species. The significance here is that the inverted hexagonal phases can most
easily be formed by weakly hydrating lipids in the absence of protein.
We have proposed a simple model (Bryant and Wolfe, 1992, Wolfe
and Bryant, 1999, 2001), which
explains the above phenomena and, so far as we are aware, all the related
data. Consider a cell whose water content has been reduced to (say) 10% by
volume. Let us suppose that a membrane rich region of the cell has this same
water content, and that its membranes are 5�nm thick. The membranes are
therefore on average 0.5�nm apart. At this separation, all hydrophilic
surfaces in water (including membranes) experience a strong repulsion
called the hydration force which decreases approximately exponentially with
separation, has a characteristic length of about 0.2�nm and whose
extrapolated value at zero repulsion (Po) is typically 10-100�MPa. Removal
of inter-membrane water in this regime either reduces the thickness of the
inter-membrane layer, and thus does considerable work against this strong
repulsion, or else reduces the area of the inter-membrane layers and thus
compresses the membranes in their plane. In practice it does both.
Figure: A cartoon of the lipid water phases of
model membranes exhibiting dehydration strains
Mechanical equilibrium in the normal direction requires that the suction in
the inter-membrane phase have the same magnitude as the repulsive force. In
the lateral direction it requires that the membranes support a compressive
stress equal in magnitude to the inter-membrane suction times the inter-
membrane separation. Severe dehydration of membrane rich regions thus
causes stacks of membranes which resemble a lamellar phase, while
compressing them laterally to make them thicker in the normal direction. In
regions of the cell rich in macromolecules, the macromolecules will also be
pushed into close separation and will suffer anisotropic internal stresses
which compress them along their longer axes, although in this case the
geometry is less simple (Wolfe and Bryant, 1999). We will concentrate only
on membranes here. Colligative properties of membranes and solutes are
compared in the figure below.
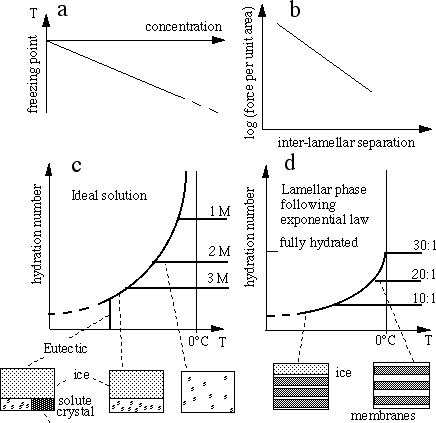
Figure: Comparing the freezing point depression
due to solutes and to membranes. (a) and (c) show the behaviour of an ideal
solution. In (a) it is shown as equilibrium freezing point depression as a
function of concentration (for sugars, the experimental curve is lower than
the line at high concentration). In (c) the same relation is represented as the
molecular ratio water:solute, as a function of temperature. The horizontal
lines show the simple fact that, for a sample with a given composition, the
composition is constant above the equilibrium freezing temperature. The
colligative effects of membranes are usually described in terms of the inter-
membrane repulsion and the inter-membrane separation. For a large range of
lipid membranes, this relation is well approximated by a repulsion that
decreases exponentially with separation, as shown in (b). Converting this to
a plot of water:lipid ratio as a function of temperature gives (d). Note the
qualitative similarity to (c). In the membrane case, the water:lipid ratio has
an upper limit: at about 30:1 the inter-membrane energy is a minimum (the
force is zero) and so adding further water to such a sample simply creates an
excess water phase (at temperatures above freezing), or more ice (at freezing
temperatures). For experimental data, see Yoon et al. (1998) or Wolfe and
Bryant (1999).
How large are these effects? Consider a single component lipid membrane.
The gel-liquid crystal phase transition involves a reduction in area per
molecule and an increase in its thickness. When these quasi two dimensional
objects are compressed in the plane by a lateral stress p, the gel phase, which has a lower area per
molecule, is favoured and the temperature of the transition is therefore
elevated due to the Clausius-Clapeyron effect. (This is the effect which, in
three dimensions, causes the boiling point of water to depend upon
atmospheric pressure.) Using typical values this yields an elevation of order
0.5��C for each extra mN/m in lateral stress. At a separation of 0.5�nm and
with a repulsion of 20�MPa, the lateral stress is about 10�mN/m (more detail
in Wolfe and Bryant, 1992; 1999).
One spectacular way in which membranes can respond to this lateral stress is
by forming non-planar geometries, including the inverted hexagonal
(HII) phase, as shown in a previous
figure. For weakly hydrating species, this geometry allows a large ratio
of lipid volume to water volume, however this transition cannot be analysed
with such a simple model because it involves energies associated with the
curvature of the interface.
In a membrane composed of mixtures of lipids having different phase
transition temperatures, a number of the phases described above can coexist
(e.g. gel-fluid coexistence) over a range of temperature. This is analogous to
solid-liquid phase coexistence between the melting points of the components
of mixtures of three dimensional materials.
In membranes at low hydration, a different mechanism can give rise to
separation into two coexisting liquid crystal phases. Consider the case where
two or more components have very different hydration properties (e.g.
different Po) - ie you have a mixture of a highly hydrating
species and a weakly hydrating species. In this case an homogeneous mix of
the components has an internal energy that is rather higher than that of the
separated phases. In many cases, this difference in internal energy is large
enough to overcome the entropy of demixing, and the mixture will phase
separate.
previous figure. Here one phase has a higher
concentration of the highly hydrating species and a higher inter-membrane
separation than the other.
This effect was first predicted theoretically (Bryant and Wolfe, 89), and then
observed experimentally using small angle X-ray diffraction and solid state
NMR (Bryant et al., 1992) for mixtures of POPC and POPE (two mixed
chain unsaturated phospholipids typical of those found in plant membranes).
In excess water, the two species are completely miscible, forming a single
lamellar phase. At 10% water content and 315�K however, the mixture
separates into two separate lamellar phases with different water separations.
Dehydration induced fluid-fluid phase separations have since been observed
for other systems (Webb et al. 1993).
The existence of fluid-fluid phase separations is not, in itself, necessarily a
danger to biological materials. However, it could be a necessary intermediate
stage in the formation of damaging inverted phases (such as
HII). Cell membranes are composed mostly of strongly
hydrating lipids that tend to form lamellar phases at all hydrations. Weakly
hydrating species (that tend to form inverted phases at low hydrations) are
normally in the minority: if they were not, then the bilayer membranes
would not be stable. Even under severe dehydrations, inverted phases are
unlikely to occur in these membranes if they remain homogeneous.
However, if fluid-fluid phase separation occurs, the weakly hydrating lipids
are concentrated into low hydration fluid phases, and are then free to
undergo the transition to an inverted phase if the hydration is low enough.
Thus fluid-fluid phase separations may be a precursor to the formation of the
inverted phases which have been correlated with membrane damage during
dehydration and freezing.
The proviso about partitioning is very important: many solutes, especially
polymers, are excluded from the inter-membrane regions. As a result, their
osmotic effect may have no direct effect on reducing the membrane stress,
and it may even increase it (Koster et al, 2000). Solutes which remain
external to the cell or to the vesicles in model systems will also give no
direct reduction to membrane stress. At temperatures above freezing, the
addition of the solute to the external solution can, in sufficiently high
concentration, dehydrate the cell or vesicle sufficiently to increase the stress.
Permeating cryoprotectants (such as DMSO) partition readily into the inter-
membrane space. In model systems, it is not simple to produce high
concentrations of non-permeating solutes in the inter-membrane space, so
comparisons among different experiments should be made carefully, unless
the inter-membrane concentration, rather than the total sample concentration,
is measured.
Yoon et al. (1998) studied lipid-solute-water systems at freezing
temperatures and used the nuclear magnetic resonance signal of the water to
determine the distribution of solute and solvent between lamellar phases and
a concentrated bulk solution phase in equilibrium with ice (they used either
D2O or deuterated solutes). For the small solute molecules
(DMSO and sorbitol) they found that the phase behaviour was close to that
expected using the effects discussed above and assuming no specific effects.
They found that the disaccharides sucrose and trehalose (which have about
twice the volume of the others) increased the hydration less than would be
expected from their osmotic effects alone. This effect was consistent with a
model in which these molecules were excluded from a very thin layer of
water closest to the lipids. Yoon et al. found that all of the solutes studied
decreased the intra-membrane stress, but that the disaccharides decreased it
more than the smaller solutes. This is only one reason why sucrose and
trehalose may occur so widely as natural cryoprotectants, however. Other
reasons concern vitrification and crystallisation.
In principle, solutes could affect the membrane stress in specific ways. If the
solutes bound to the membrane surface, for example, one would expect a
modification in the hydration force. One study of inter-membrane forces
using the Surface Forces Apparatus found no specific effects on the inter-
membrane force due to DMSO, sorbitol or trehalose, however this study was
limited for technical reasons to concentrations of about 1�kmol.m-
3 (Pincet et al., 1994).
In practice, the measured transition temperature in the presence of a glass is
in the range 10-60�C below the fully hydrated transition temperature
To. Koster et al. (2000) have measured the mechanical
properties of a relevant sugar glass, and found Young's modulus to be about
20 GPa. Using parameters for DPPC (Guldbrand et al., 1982), and eq. (1),
the compressive stress generated if the lipid remains fluid 20�C below
To is about 40�mN/m, which corresponds to a strain of about
0.4% in the glass. That level of strain is easily supportable by a solid.
The importance of this effect is clear: if the solution is vitrified while
the lipids are in the liquid crystal phase, then the transition temperature will
be lowered dramatically, and the membranes will remain in the fluid state. In
addition, the formation of the glass has two other important effects. First, if
the solution vitrifies then further dehydration will be extremely limited; and
second, if the sample is vitrified, solute crystallisation will be restricted. This
is beneficial as the protective effects of solutes mentioned in the previous
sections can only occur if the solutes remain in solution.
A list of publications by Joe Wolfe.
Avoiding intracellular ice formation
Supercooling
Supercooling refers to taking a liquid below its equilibrium freezing point,
without freezing. Biological solutions in situ can usually supercool a
couple of �C or more. From the point of view of the organism, this freezing
avoidance mechanism has the advantage that the solutions remain liquid and
allow relatively normal, though slower, metabolism. It has the disadvantage
that a supercooled solution is unstable: if an ice crystal is introduced, it tends
to freeze immediately, forming ice and a more concentrated solution. A very
small volume of a pure solution may supercool as much tens of �C, but
biological solutions are threatened by ice nucleators which may initiate
freezing either inside or outside the cell. Supercooling is a vital strategy for
some Antarctic fish that live in a solution with a higher concentration and
greater freezing point depression than those of their own tissues. Their blood
carries a potent protein "antifreeze". This substance, present only in tiny
molar concentrations, does not depress the equilibrium freezing temperature,
but works by impeding the growth of ice crystals (DeVries, 1984). The
environmental temperature for such fish has a robust lower bound: they will
not encounter temperatures colder than the freezing temperature of the
ocean. Without such a lower bound, the supercooling strategy is a dangerous
one. Plant leaves often supercool a few degrees and may thus survive mild
frosts without freezing damage, whereas slightly colder temperatures can
cause extensive damage (Lutze et al., 1998). The importance of supercooling
in cryopreservation is that it allows vitrification, which we look at later.Freezing point depression
Many plants and animals found in cold environments accumulate soluble
molecules (solutes) in their intracellular and often extracellular
solutions (Leopold, 1986, Lee, 1989, Koster and Lynch, 1992). These
solutes lower the equilibrium freezing temperature. Another result is that the
organism is more resistant to dehydration, for osmotic reasons discussed
below. Freezing point depression of more than a few �C requires rather
highly concentrated solutions (see the conversion chart below). Large
increases in the concentration of salts are rare in biological systems for the
electrical reasons mentioned above. Solutes which can be tolerated in high
concentrations (compatible solutes) include a number of sugars.
High concentrations of such molecules increase the viscosity and thus reduce
diffusion in solutions. This slows metabolism, but has advantages for
slowing further dehydration and for vitrification, which we discuss later. The
main importance of freezing point depression in cryopreservation is in the
avoidance of crystallisation in the high-temperature, low viscosity
r�gime.Dehydration
Freezing usually occurs outside the cell first, because that's where there are
more freezing nuclei, and because the extra-cellular solution has a larger
volume than the intracellular solution. When this happens, the extra-cellular
solutes are concentrated in a small quantity of unfrozen water, which
necessarily has a higher osmotic pressure. This causes water to leave the
cell. The characteristic time for water to flow out of cells under these
conditions is tens of seconds (Wolfe and Bryant, 1992), so we refer to
cooling as fast or slow according to whether substantial temperature changes
are possible over this time scale. Cryopreservation usually uses fast cooling
and so, when and if extracellular ice occurs, cells do not have time to
dehydrate severely, although there is usually some dehydration of
cryobiological importance. Environmental cooling, on the other hand, is
slow so water is usually close to equilibrium, except under extreme
dehydration. 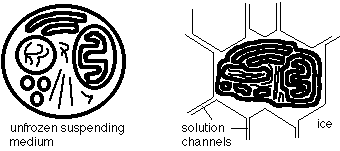

Intracellular vitrification
In cryopreservation, the usual goal is to achieve intracellular vitrification
while avoiding intracellular ice formation and membrane damage. One of
the deciding factors is the cooling rate. If a liquid is cooled sufficiently
quickly, it can avoid freezing and vitrify (form an amorphous, glass phase).
The necessary cooling rates are extremely high for pure liquids (e.g. 10
million degrees per second for pure water), but much more realistic for
solutions. For aqueous solutions of typical cryoprotectants, cooling rates of
about 0.1-10��C/s (roughly 10-1,000 �C/min) are sufficient to achieve
vitrification.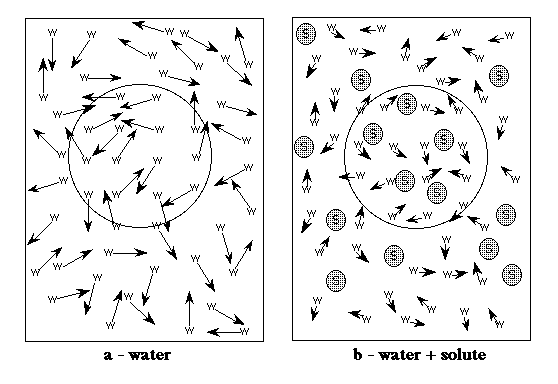
Fast cooling and warming
So, how to get cells past the dangers that await them in freezing and
warming? The next figure shows how for the case of fast cooling and
warming---i.e. in artificial cryopreservation. In this case, the degree of
osmotic contraction is usually small, as discussed above. Osmotic
contraction depends on cooling rate (Mazur, 1963), which is in part
responsible for there being an optimum in cooling rate. At very slow cooling
rates, substantial osmotic contraction occurs, and this may be fatal in itself
(discussed later). At very high rates, there is little osmotic contraction, so the
solute concentration in the cytoplasm remains low. This makes vitrification
less likely, and intracellular freezing more likely. At moderate rates (the
value depends on the osmotic equilibration time of the cell and the
propensity of its cytoplasm to nucleate ice), some non-fatal contraction
which causes the concentration to increase sufficiently that vitrification can
occur. A moderate osmotic contraction can be controlled by means other
than cooling rate: one method is to cool the cells to a relatively high freezing
temperature and allow them to contract to equilibrium prior to fast cooling.
Another is to add a non-penetrating cryoprotectant, i.e. an
extracellular solute which increases the extracellular osmotic pressure.
Hydroxyethyl starch (HES) or dextran are used. This has two effects: the
moderately contracted cells avoid IIF during the initial cooling; and the
extracellular solution vitrifies at relatively high temperatures. The detailed
mechanisms of polymer cryoprotection are still unclear (e.g. Bryant et al.,
94). 
Slow cooling and warming
Even at sunrise and sunset, cooling and warming rates in nature are usually
low compared to the characteristic time for osmotic equilibration. As a
result, extracellular freezing often causes dehydration which approaches
hydraulic equilibrium. An exception occurs when an intracellular solution
vitrifies and thereafter undergoes no or very little further loss of water. In
some cases of dehydration without vitrification, the viscosity may still be
large enough to prevent or to slow further dehydration. The next Figure
shows a flow chart for slow cooling and warming.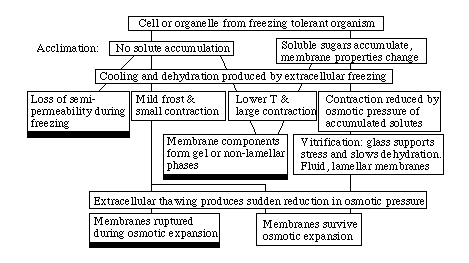
Rupture during freezing and contraction
During this process of freezing and osmotic contraction, some cells rupture.
Simple mechanical rupture by the advancing ice crystals is one likely cause.
Another cause of rupture may be electrical: the large transient electric field
associated with an advancing ice front in a weak electrolyte solution causes a
potential difference across the cells, which is large enough to rupture
membranes, and which is correlated with rupture (Steponkus et al.,
1985).Irreversible osmotic contraction
Osmotic contraction is often irreversible. Cells isolated from the leaves of
some frost sensitive plant species have a limited ability to contract and to re-
expand osmotically, and this is correlated with the occurrence of freezing
damage. Plants which have been acclimated to low (but not freezing)
temperatures are much less susceptible to frost damage at modest freezing
temperatures, and their protoplasts (isolated cells) are capable of
much larger osmotic excursions (Wiest and Steponkus, 1978). Protoplasts
have the property that they become spherical when suspended in media
having a large range of osmotic pressures. When the concentration of the
suspending medium is increased, (the osmotic equivalent of extracellular
freezing) the protoplasts initially become flaccid, but over several minutes
they become spherical with a smaller area. The plasma membrane has a
small resting tension---a few tenths of a mN/m. This is the process which is
not always reversible. When the external medium is abruptly diluted back to
its original composition (the osmotic equivalent of thawing), the volume
increases rapidly and the area increases almost equally rapidly. The plasma
membrane can support a tension of only several mN/m without rupture.
Substituting this and a typical radius (10 microns) the Young-Laplace
relation gives about 1 kPa as the maximum hydrostatic pressure that can be
supported. This is about a thousand times less than the osmotic pressures
involved, and so may be neglected for such cells (though not necessarily for
small robust cells in less concentrated media, such as red blood cells). It
follows that the area of the cell is determined by the flux of water into the
cell. For a halving of the concentration of the suspending medium, an area
increase of order 50% is required. The area elastic modulus for such a
membrane is about 200�mN/m, so it is capable of an elastic expansion of
only a few percent without rupture. Once the membrane stretches and its
tension increases, membrane material is incorporated into the membrane at a
rate which is a strong function of the applied tension (Wolfe and Steponkus,
1983). The biophysics of this problem is attractive in that all of the relevant
parameters and functions can be measured: the osmotic properties of the
solution, the hydraulic conductivity of the membrane, its relevant elastic
modulus, the rate of membrane material incorporation and the probability of
rupture as functions of tension. Further, the differential equations for tension
and probability of lysis have analytic, though awkward, solutions (for
details, see Wolfe et al., 1985, 1986, Dowgert et al., 1987). One result is
unexpected: membranes from acclimated plants are slightly less robust than
those from non-acclimated. However, they more than compensate for this by
having a much greater rate of incorporation of new material.Damage produced by severe dehydration
Freezing damage to frost tolerant plants is correlated with a different cellular
malfunction: loss of membrane semipermeability in the freezing-induced
dehydrated state. Like the irreversible osmotic contraction described above,
this symptom can be reproduced by osmotic manipulation at room
temperature: protoplasts dehydrated in high concentration suspending media
dehydrate but, when the medium is diluted, they fail to expand osmotically.
A range of desiccation tolerant species of plants and animals also support
substantial dehydrations but, in some cases, dehydration below a critical
value (of order 10% water content) can be fatal. In this case, freezing
damage and desiccation damage are similar and cryobiology and
anhydrobiology overlap. In both cases, accumulation of solutes can reduce
or prevent damage in model systems and species that are freezing or
desiccation tolerant are observed to accumulate solutes, especially sucrose
and trehalose (Leopold, 1986).
The effect of solutes (cryoprotectants)
The presence of high concentrations of low molecular weight solutes in
model membrane systems reduces the incidence of the effects associated
with dehydration damage: they reduce the elevation of the membrane
transition temperature and they reduce the occurrence of non-lamellar
phases. This may be one reason why freeze- and desiccation- adapted
species have evolved to accumulate solutes, often sucrose, trehalose or other
sugars (eg Leopold, 1986, Crowe et al., 1988).
Osmotic effects
At a given chemical potential of water, the presence of more solutes requires
the presence of more water (see first figure above). A cell or a vesicle that
has a higher internal concentration at temperatures above freezing will
contract less in equilibrium with ice at any given freezing temperature.
Further, the addition of any new solute requires a reduction in the
concentration of others already present. The presence of a high concentration
of sugars reduces the concentration of ions that is required to produce a
given osmotic pressure. So the presence of sugars reduces the dangerous
high ion concentrations mentioned earlier. Solutes which can be
accumulated in large concentrations without producing toxic effects of their
own are called compatible solutes (Brown, 1976).
Reduction in mechanical stress
Provided that the solutes partition into the inter-membrane layers, their
presence contributes osmotically to the lowering of the chemical potential of
water. The larger the osmotic term, the smaller the suction and so the lower
the stress imposed on the membrane. This effect can be considerable (Yoon
et al, 1998; Wolfe and Bryant, 1999).Vitrification and ultrastructure
An important extra consideration arises when and if the inter-membrane
layer vitrifies. It has been found that under these conditions the dehydration-
induced increase in the transition temperature is dramatically reduced, and
the transition temperature can be reduced below the fully hydrated value
To (Koster et al., 1994, Crowe et al., 1998). Consider a
membrane at a fixed hydration at a temperature a few degrees above its
transition temperature Tm. As the temperature is lowered to
Tm the transition takes place, accompanied by a reduction in
area per lipid (see previous figure). If however the
inter-membrane aqueous solution is vitrified, the transition can not take
place. The vitrified layer is a solid, so is capable of supporting considerable
mechanical stress. If the temperature is lowered through Tm,
the glass will impede the reduction in area necessary for the gel phase to
form. As the temperature is lowered below Tm, the glass will
support an increasing compressive stress in the membrane. At some point
below To, the stress will become large enough to overcome the
presence of the glassy matrix, and the gel transition will occur. During
warming, the transition temperature will remain the same. Conversely, if the
membrane is in the gel phase when vitrification occurs, the transition
temperature will be raised above the fully hydrated transition temperature
To, during both cooling and warming (Zhang and Steponkus,
1996).References and links
Much of the above essay comes from the paper
Other parts come from some of our other publications, also available on line:
Related links
Joe
Wolfe
/ J.Wolfe@unsw.edu.au
/61-2-9385 4954 (UT+10,+11 Oct-Mar)
Gary Bryant / Gary.Bryant@rmit.edu.au
/61-3-9925-2139 (UT+10,+11 Oct-Mar)
