| |
||
| |
||
| |
||
| |
|
|
|
|
| The cochlea of the normal inner ear transforms an input mechanical vibration
(essentially a filtered version of the acoustic signal input to the ear)
to action potentials in the fibres of the auditory nerve. The cochlear
is a rigid, coiled tube, divided mechanically into two along its length
by the basilar membrane. The small bones of the middle ear input a displacement
signal to one side of the tube via a window. This signal drives a transverse
wave in the basilar membrane, whose cutoff frequency decreases along its
length. As a result, high frequencies cause maximum vibration at the window
end, and low frequencies cause maximum vibration at the other. In the
normal ear, action potentials are produced in an array of hair cells which
reside on the basilar membrane. Ohm [2] and Helmholtz [3] proposed that
pitch was encoded tonotopically, i.e. by the place along the basilar membrane
of the nerve stimulated (place theory). Seebeck [4] argued that nerve
pulses were produced by each vibration and that their rate determined
the perceived pitch (rate theory). In the place theory, it is difficult
to explain the observed fine resolution of frequency (~0.2%). The rate
theory cannot readily explain the perception of tones with frequencies
many times greater than the maximum firing rate of neurones. Despite many
elegant acoustic experiments, the relative importance of rate and place
are still debated because, in the normal ear, the rate of mechanical stimulation
of the basilar membrane is strongly correlated with position. Cochlear
implants allow the local electrical stimulation of different regions of
the cochlea at different rates. A range of experiments have studied pitch
using CIs: Simmons et al [5] reported pitch estimates from a single subject
with low resolution in position. Pitch as a function of stimulation rate
was reported by Pijl [6] and by Collins et al [7].
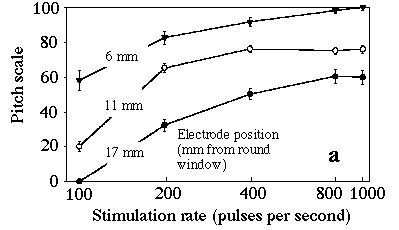
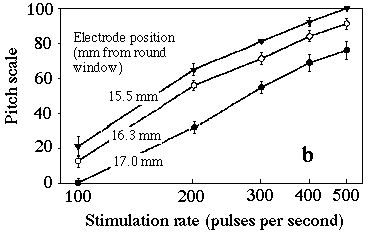
Our study extends the work by these researchers and uses the method of pitch scaling [7,8] which has the advantages that it does not require matching of percepts that may differ in several different perceptual parameters, and that it can readily be understood and used by subjects with little knowledge of music. We studied six volunteers with implants which allowed fine resolution in rate and place, and we present perceived pitch as a function of rate and place of stimulation. The results show remarkable consistency, given the subjective nature of the test. Six adults who had lost their hearing at ages between 5 and 45 years volunteered for this study, which is part of a project to improve the performance of CIs in delivering perception and appreciation of music. The stimuli were 1.00 s pulse trains of biphasic rectangular pulses: a 100 microsecond pulse, a 25 microsecond gap then a 100 microsecond pulse in the opposite direction. The stimuli were loudness balanced by asking each subject first to increase the control of the current level to achieve a "medium-loud" level, then to compare all stimuli in turn with the middle rate, middle position stimulus until the subject was satisfied with loudness equivalence. Figure 2a shows the result for the experiment over the larger range. At low frequencies, the pitch is strongly dependent on both rate and place but, at rates above several hundred pps, the stimulation rate has little effect and pitch decreases with distance from the round window. Figure 2. Average of the scaled pitch estimate (plus or minus s.e.) as a function of stimulation rate and electrode position. Higher number electrodes are inserted further into the cochlea (most distant from the window). The different electrode pairs in (b) are displaced from each other by 0.75 mm. To elicit reports of perceived pitch from a subjects unfamiliar with the names of musical intervals, the pitch scaling method [7,8] was used: subjects were asked to rate pitch on an arbitrary scale from 0 (low pitch) to 100 (high). Seven examples of each stimulus were delivered and evaluated. Presentation order was random. For each subject in each of the two experiments, the responses were scaled as a percentage of the total range used by the subject. The number, time and good will of volunteers are generous but finite. This limits the volume of parameter space that may be investigated. One experiment investigated rates from 100 to 500 pps, applied between the three pairs of electrodes at the end of the array most distant from the round window (1b). The positions given are the average values measured in another study [9]. Five of the subjects returned for another experiment in which rates between 100 and 1000 pps were applied to three pairs of electrodes widely spaced along the array (1a). All used Nucleus (TM) CI22M implants and either SPECTRA-22(TM) or SPrint(TM) processors programmed with the SPEAKTM coding strategy (Cochlear Ltd.). All subjects normally use biphasic pulses applied between pairs of electrodes separated by one temporarily inactive electrode; that stimulation mode was used in this study. Figure 2b shows the average of the scaled pitch for all subjects for the experiment with smaller rate and place range. The difference between electrodes at 15.5 mm and 16.3 mm is significant at 0.05, which suggests that the resolution of position in this context is of the order of, or less than, one electrode spacing (0.75 mm). The logarithmic dependence of pitch on rate invites comparison with normal hearing, where notes in the equal tempered chromatic scale of Western music are equally spaced on a log frequency scale. For the CI subjects, pitch also depends on place of stimulation, decreasing with distance from the round window. This can be compared with the tonotopic arrangement of the normal ear where a doubling in the frequency of the acoustic signal corresponded to a displacement of about 4 mm along the basilar membrane for frequencies above several hundred Hz, and smaller displacements for lower frequencies [10]. Because the pitch scales shown in Fig 2b are approximately logarithmically dependent on rate, we can calculate that a doubling in stimulation rate corresponds to a displacement of about 2 mm in this range. For the series of experiments reported in Fig 2a, the displacement corresponding to a doubling of stimulation rate depends on position and rate. It is about 4-6 mm at low rates and decreases for higher rates. The results for electrodes at 17 mm are slightly different between the two experiments. This is partly because the pitch scale is arbitrary and the experiments were conducted at different times. It is also possible that the task of assigning pitch is more difficult over a much larger range of the parameters. The apparent saturation of the dependence of pitch on stimulation rate is not surprising at rates which are greater than the maximum firing rate of neurones. These results may not simply be compared with normal hearing, however, because the differential mechanical stimulation of hair cells is rather different from the electrical stimulation by the CI of many or all of the cells between or near the two electrodes. The influence of rate and place on pitch perception for these post-lingually deafened subjects nevertheless suggests that both rate and place are important in pitch coding for normal hearing at low frequencies, but that place alone dominates at sufficiently high frequencies. Acknowledgements. RF was supported by an Australian Postgraduate Award (Industry). We thank Stephanie Shaw and our volunteer subjects. References 1 Békésy, G. von (1960) "Experiments in Hearing", McGraw-Hill, New
York. |
|
|
|
|
|
|